






Tous les moyens de contacter une animatrice LLL, par téléphone, par mail, en réunion...


Savez-vous que LLL France fonctionne grâce à vos cotisations et à vos dons ?


Vous trouverez ici les dernières études parues sur l'impact de l'allaitement pour l'enfant et la mère, ainsi que des témoignages de mères

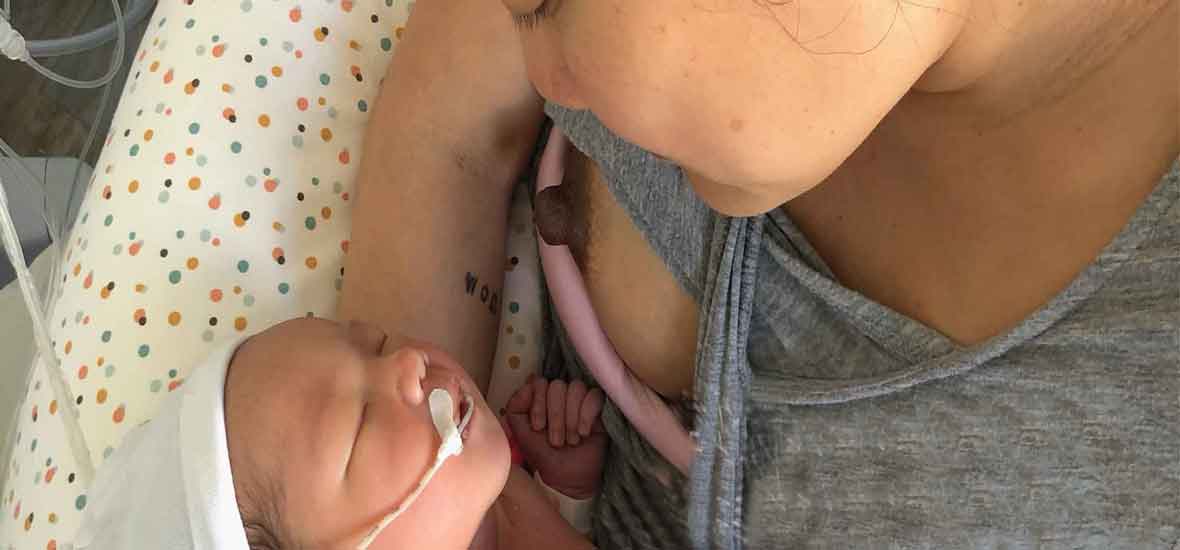
Avec un soutien personnalisé, des informations actualisées, adaptées, il est bien souvent possible de dépasser les difficultés des premiers jours, des premières semaines

Semaine mondiale de l'allaitement maternel 2023

La certification Qualiopi a été délivrée à LLL France au titre de la catégorie d'actions suivante : actions de formation